
Kritik an derPolyvagaltheorie:Eine umfassendeAnalyse
vonStephen W. Porges, PhD
Renommierter Universitätswissenschaftler,
Kinsey Institute, Indiana University
Überblick
Die Polyvagal-Theorie hat einen biologisch fundierten Rahmen zum Verständnis von autonomer Regulation, Sicherheit und sozialer Interaktion geschaffen und damit die Forschung und Anwendung in den Bereichen Neurowissenschaften, Psychologie, Medizin und Pädagogik beeinflusst. Mit ihrem Bekanntheitsgrad haben auch die kritischen Stimmen in akademischen Kreisen und öffentlichen Foren zugenommen. Diese Seite bietet eine prägnante, evidenzbasierte Erläuterung der Kernaussagen, des Anwendungsbereichs und der wissenschaftlichen Grundlagen der Polyvagal-Theorie mit dem Ziel, durch Verweise auf Primärquellen und den aktuellen Stand der Neurophysiologie eine zutreffende Einschätzung zu unterstützen.
Die veröffentlichten Kritiken an der Polyvagal-Theorie, insbesondere die von Paul Grossman und E. W. Taylor, weisen ein wiederkehrendes Muster von Fehlinterpretationen und sachlichen Fehlern auf. Trotz mehrfacher Klarstellungen in der Peer-Review- geprüften Fachliteratur (Porges, 2007b, 2022, 2023, 2025a, 2025b) führen diese Kritiken noch immer zu konzeptionellen Fehlern sowohl im akademischen Diskurs als auch in der klinischen Anwendung. Grossmans Kritik von 2023 greift Punkte auf, die zuerst von Taylor vorgebracht wurden und stützt sich dabei stark auf dessen Fehlinterpretationen anatomischer und evolutionärer Fakten, die rhetorisch als Gewissheiten formuliert werden und Leser, die mit der Basisliteratur hinter der Theorie nicht vertraut sind, in die Irre führen können.
Neben Peer-Review-geprüften Publikationen haben auch öffentlich zugängliche Quellen – insbesondere der Wikipedia-Eintrag zur Polyvagal-Theorie – zur Verbreitung fehlerhafter Darstellungen beigetragen. Dieser Eintrag beruft sich vor allem auf zwei Kritiken: Neuhuber und Berthoud (2022) sowie Doody, Burghardt und Dinets (2023). Zwar handelt es sich bei beidem um per Peer-Review geprüfte Artikel, ihr Erscheinen in einer von Grossman herausgegebenen Sonderausgabe von Biological Psychology jedoch deutet auf eine bestimmte redaktionelle Ausrichtung hin, die wahrscheinlich sowohl thematisch als auch konzeptionell ihren Betrachtungswinkel geprägt hat.
Die Kritik von Neuhuber und Berthoud ist beispielhaft für eine anatomisch-funktionelle Fehlinterpretation. Sie stellen zwar die von der Polyvagal-Theorie postulierte funktionelle Interpretation der Vagusbahnen in Frage, bestreiten dabei aber weder die zentralen neuroanatomischen Unterschiede – insbesondere die Trennung zwischen dem dorsalen motorischen Vaguskern (DMNX) und dem Nucleus ambiguus (NAmb) – noch die voneinander abweichende Myelinisierung ihrer Efferenzen. Sie lassen jedoch einen wichtigen Lehrsatz der Theorie außer Acht: dass das Social Engagement System (SES) aus Strukturen der embryonalen Pharyngealbögen hervorgeht und dass die Kommunikation zwischen dem kardioinhibitorischen Vagus und dem SES weitgehend indirekt erfolgt, mit Ausnahme der direkten Bahnen, an denen der Trigeminusnerv beteiligt ist. Wikipedia verzerrt diese Kritik bei seiner Zusammenfassung noch weiter, indem es sie als kategorische Widerlegung der anatomischen Grundlage der Polyvagal-Theorie darstellt, was der ursprüngliche Artikel gar nicht behauptet.
In ähnlicher Weise stellt die Kritik von Doody, Burghardt und Dinets den phylogenetischen Rahmen der Theorie falsch dar. Sie bestreiten das, was sie als starre Zweiteilung zwischen „asozialen Reptilien” und „sozialen Säugetieren” interpretieren. Die Polyvagal-Theorie ist jedoch ausdrücklich auf Säugetiere ausgerichtet und definiert soziales Verhalten aus der Perspektive von Säugetieren. Sie betont, dass Sozialität im Kontext der Polyvagal-Theorie in einer säugetierspezifischen autonomen Innovation verwurzelt ist, dem ventralen Vaguskomplex, der für die Regulation des Social Engagement Systems verantwortlich ist. Reptilien mögen zwar adaptive soziale Verhaltensweisen zeigen, diese Verhaltensweisen werden jedoch durch andere Signalsysteme und neuronale Architekturen vermittelt und erfüllen andere evolutionäre Funktionen. Das Sozialverhalten von Reptilien stellt zwar eine evolutionäre Adaption dar, entspricht jedoch nicht der Sozialität von Säugetieren gemäß ihrer Definition in der Polyvagal-Theorie. Durch die Vermischung verhaltensmäßiger Analogien mit neurophysiologischer Homologie übersieht die Kritik den domänenspezifischen Anwendungsbereich der Theorie. Für die Öffentlichkeit bestimmte Zusammenfassungen, darunter auch die bei Wikipedia, verstärken diese Fehlinterpretation noch, indem sie die Kritik überbewerten und den Fokus der Polyvagal-Theorie auf die hierarchische autonome Evolution ignorieren.
Entscheidend ist, dass das Problem nicht nur darin besteht, dass diese Artikel existieren, sondern dass sie von Seiten einer öffentliche Plattform so gefiltert wurden, dass ihr Inhalt überspitzt wie auch falsch dargestellt wird. Diese Fehldarstellung auf mehreren Ebenen – Ergebnis persönlicher Voreingenommenheit und nicht von sorgfältiger wissenschaftlicher Arbeit – trägt zur Verbreitung irreführender Informationen bei. Sie unterstreicht die Notwendigkeit einer wissenschaftlichen Bewertung, die auf der direkten Auseinandersetzung mit Primärquellen basiert und nicht auf Interpretationen, die von der redaktionellen Ausrichtung, selektiven Hervorhebungen oder sekundär Verbreitetem geprägt sind.
Um die Natur dieser Fehlinterpretationen zu verdeutlichen, benennt Tabelle 1 die wiederkehrenden Fehlerdomänen in der Kritik an der Polyvagal-Theorie und skizziert, wie diese sich manifestieren und welche Konsequenzen sie für eine korrekte theoretische Interpretation haben.
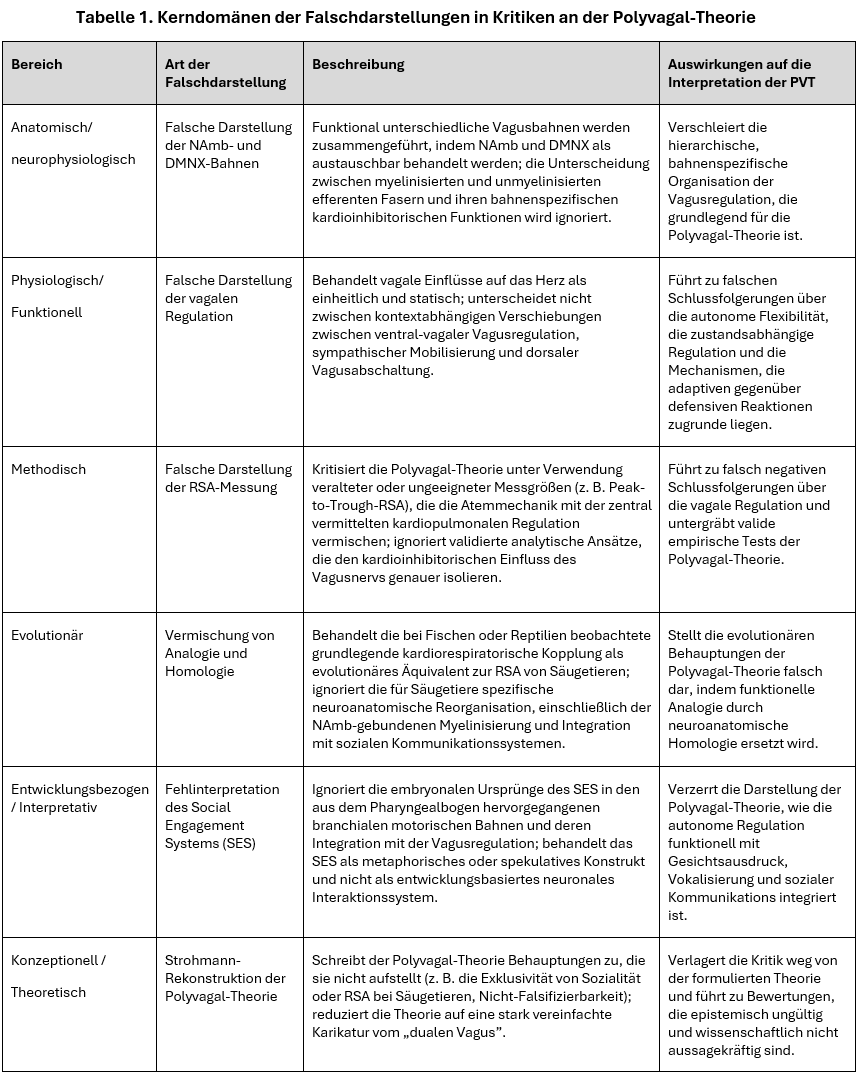
Wie in Tabelle 1 dargestellt, umfassen diese Bereiche anatomische, physiologische, evolutionäre, konzeptionelle und methodologische Dimensionen. Der konkrete Inhalt der Kritiken variiert zwar, was sich jedoch eint, ist die Fehlertypologie – sie untergraben die Kohärenz und wissenschaftliche Validität der Polyvagal-Theorie durch falsche Darstellung von Kernkonzepten oder Anwendung fehlerhafte Bewertungskriterien. Der hier vorgelegte strukturierte Rahmen bietet ein diagnostisches Prisma, durch das sich die epistemische Integrität sowohl akademischer als auch öffentlicher Kritik bewerten lässt.
Während Tabelle 1 generell wiederkehrende Fehlerbereiche in den Kritiken identifiziert, ordnet Tabelle 2 diese Verzerrungen konkret wichtigen Veröffentlichungen zu. Dabei wird beleuchtet, wie Taylors grundlegende Falschdarstellungen Muster begründete, die in späteren Kritiken reproduziert und erweitert wurden, insbesondere in denen von Grossman. Zitate von Taylor und seinen Kollegen (z. B. Campbell et al., 2006; Monteiro et al., 2018; Sanches et al., 2019; Taylor et al., 2022) weisen auf wiederkehrende Muster in verschiedenen Publikationen hin; spezifische vorherrschende Fehlertypen in Bezug auf einzelne Artikel werden in einer späteren typologischen Zusammenfassung (siehe Tabelle 4) und in der kommentierten Bibliografie aufgeführt.
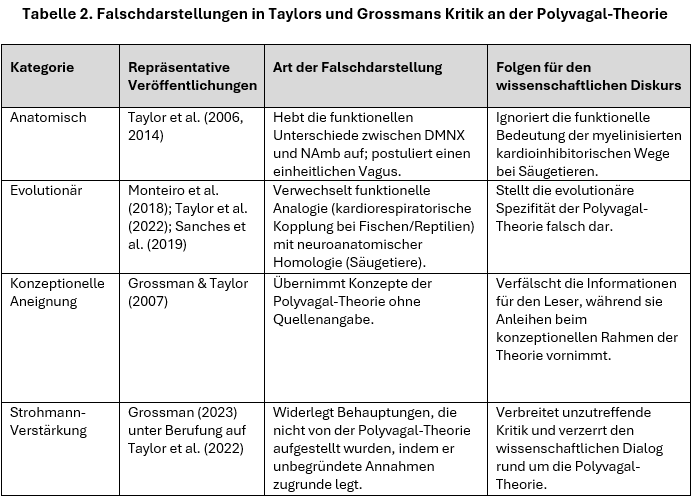
Wie in Tabelle 2 dargestellt, vernachlässigt Taylor in seinen Veröffentlichungen wiederholt anatomische Unterschiede und verwechselt phylogenetische Konstrukte, die für die Polyvagal-Theorie von zentraler Bedeutung sind. Grossmans spätere Schriften verstärken diese Fehlinterpretationen und betten sie in Kritiken ein, die rhetorisch als Gewissheiten formuliert werden, empirisch aber falsch sind. Diese Weitergabe von Fehlern trägt zu einer vielschichtigen Fehldarstellung der Polyvagal-Theorie sowohl in akademischen Kreisen als auch gegenüber der Öffentlichkeit bei.
Während Tabelle 1 und 2 allgemeine und grundlegende Fehlinterpretationen kategorisieren, befasst sich Tabelle 3 direkt mit den wichtigsten Kritikpunkten von Grossman bewertet sie anhand etablierter empirischer und neurophysiologischer Belege.
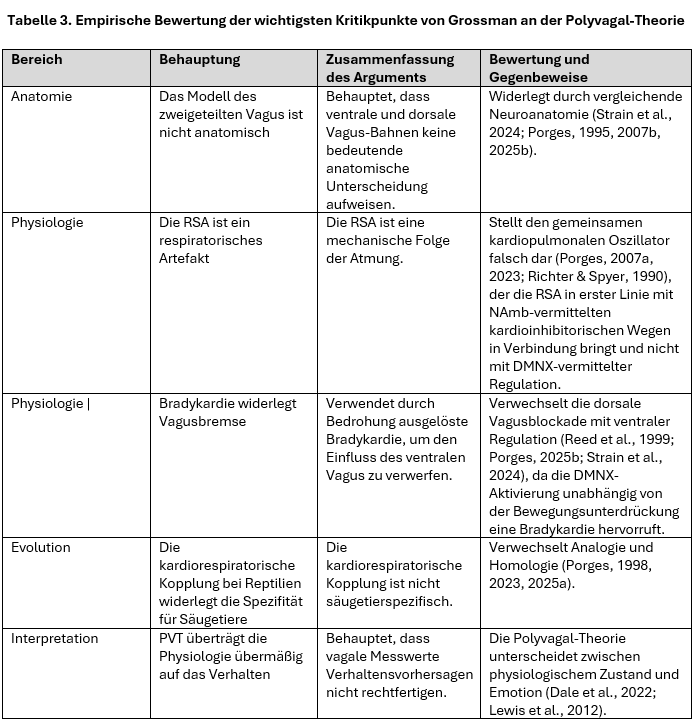
Tabelle 3 fasst die wichtigsten Behauptungen von Grossman (2007, 2023) zusammen und bewertet sie anhand aktueller empirischer und neurophysiologischer Erkenntnisse. Ob auf dem Gebiet der Anatomie, Physiologie, Evolution und Interpretation – in diesen Kritikpunkten schlagen sich durchweg eher Missverständnisse als empirisch fundierte Meinungsverschiedenheiten nieder – was die Notwendigkeit einer auf der Primärliteratur basierenden Bewertung unterstreicht.
Tabelle 3 dient nicht nur als Zusammenfassung der neurowissenschaftlichen Beweise, die Grossmans Kritik widerlegen, sondern auch als Demonstration der inneren Kohärenz der Polyvagal-Theorie. Durch ihre Übereinstimmung mit aktuellen Erkenntnissen aus den Bereichen Neuroanatomie, Physiologie, Entwicklungspsychobiologie und Evolutionsbiologie bewahrt die Theorie ihre konzeptionelle Integrität und interdisziplinäre Konsistenz und unterstreicht damit die Robustheit ihrer Erklärungen statt einer Angreifbarkeit durch reduktionistische Kritik.
Alles in allem sagen die hier angesprochenen Kritikpunkte – ob anatomischer, physiologischer, evolutionärer oder konzeptioneller Natur – mehr über die Interpretationsweise ihrer Urheber aus als über die Gültigkeit der Polyvagal Theorie selbst. Die sich hartnäckig haltende Fehlinterpretation, insbesondere in öffentlich zugänglichen Zusammenfassungen, unterstreicht die Notwendigkeit, bei der Bewertung der Behauptungen der Polyvagal-Theorie auf die Primärquellen zurückzugreifen. Wie diese Analyse zeigt, bleibt die Polyvagal-Theorie ein kohärentes und empirisch fundiertes Rahmenwerk, dessen interdisziplinäre Reichweite sowohl die Grundlagenforschung in den Neurowissenschaften als auch translationale Anwendungen weiterhin beeinflusst.
Methodische Falschdarstellung und die Behauptung zum RSA-Artefakt
Grossmans wiederholte Behauptung, dass die RSA lediglich ein Atemartefakt sei, beruht auf seiner Verwendung der Peak-to- Trough-Methode zur Quantifizierung der RSA – einer Technik, deren methodische Mängel schon seit langem anerkannt sind. Diese Methode, die zuerst von Byrne & Porges (1993) kritisiert und später von Lewis et al. (2012) bestätigt wurde, verstößt gegen wichtige statistische Annahmen, reagiert übermäßig empfindlich auf Atmung und Signaltrends und besteht die Tests auf Normalverteilung und Stationarität nicht. Die Porges-Bohrer- Methode hingegen reagiert besser auf vagale Blockaden, wird weniger von respiratorischen Variablen beeinflusst und eignet sich besser für eine valide parametrische Analyse.
Trotz langjähriger Kritik an diesem Vorgehen wurde der Peak-to- Trough-Ansatz von Grossman weiterhin angewendet, was zu einer systematischen Verzerrung und einer irreführenden Interpretation der Beziehung zwischen der RSA und dem ventralen Vagustonus geführt hat. Seine Weigerung, die Analysemethoden anzupassen oder empirische Widerlegungen anzuerkennen, hat dazu geführt, dass dieser Fehler über mehr als zwei Jahrzehnte hinweg in Veröffentlichungen fortbesteht.
Infolgedessen basieren viele der veröffentlichten Kritiken an der Polyvagal-Theorie nicht auf gegenteiligen Nachweisen, sondern auf einem einzigen, nicht korrigierten methodischen Fehler. Die Behebung solcher Fehler ist entscheidend für die Wiederherstellung der wissenschaftlichen Präzision und analytischen Nachvollziehbarkeit in Diskurs um die Polyvagal-Theorie.
Kritisiert Grossman die Polyvagal- Theorie oder eine Falschdarstellung von ihr?
Eine genaue Analyse von Grossmans Kritik (2007, 2023) zeigt, dass seine Argumente häufig auf eine Version der Polyvagal- Theorie Bezug nehmen, deren Formulierung von der in der Primärliteratur verwendeten abweicht. Diese Abweichung entsteht durch selektive Zitate, das Zurückgreifen auf veraltete Modelle der Vagusphysiologie und die Fehlinterpretation vergleichender neuroanatomischer Beweise. Infolgedessen werden der Polyvagal-Theorie Prämissen zugeschrieben, die sie nicht vertritt (z. B. dass die RSA nichts mit der Atmung zu tun hätte oder dass NAmb nur bei Säugetieren vorkäme), und diese ihr zugeschriebenen Positionen werden anschließend abgelehnt.
Dieses Muster spiegelt eher ein kategorisches Missverständnis wider als eine Meinungsverschiedenheit, die auf konkurrierenden empirischen Daten beruht. Insbesondere verwechselt Grossman wiederholt funktionelle Analogie mit neuroanatomischer Homologie und übersieht dabei die phylogenetische Begründung, die in der Polyvagal-Theorie dargelegt wird. In ähnlicher Weise verschleiert die Nichtunterscheidung zwischen DMNX und NAmb die anatomische Grundlage des hierarchischen Modells der autonomen Regulation in dieser Theorie.
Obwohl die grundlegenden Arbeiten zur Polyvagal-Theorie in diesen Kritiken häufig zitiert werden, stehen die kritisierten Punkte in keinem Zusammenhang mit einer korrekten Darstellung der ursprünglichen neurophysiologischen Behauptungen der Theorie. Stattdessen stützen sich Grossmans Kommentare auf eine stark isolierte Betrachtung spezifischer Teilbereiche der Neurowissenschaften – wie vergleichende Anatomie oder
Neurophysiologie –, ohne integrative theoretische Modelle zu berücksichtigen, die Struktur, Funktion, Entwicklung und Verhalten zueinander in Beziehung setzen. Infolgedessen konzentrieren sich die Kritiken unverhältnismäßig stark auf sekundäre Interpretationen und nachgelagerte klinische Anwendungen und weniger auf die in der grundlegenden Literatur formulierten Kernkonzepte. Diese als methodologische Kritik formulierten Arbeiten perpetuieren die ursprünglich von Taylor und Kollegen eingeführten Falschdarstellungen, indem sie Mechanismen durch Metaphern ersetzen und eine Version der Polyvagal-Theorie bewerten, die in den Primärquellen nie vertreten wurde.
Ethische Implikationen
Falsche Darstellungen der Polyvagal-Theorie haben epistemischen Schaden angerichtet – sie verwirren Kliniker, führen Lehrende in die Irre und untergraben die translatorische Klarheit. Diese Folgen sind
nicht nur akademischer Natur, sondern behindern auch die korrekte Anwendung einer Theorie, die auf validierter Neuroanatomie, robusten physiologischen Messwerten und einem kohärenten neuroethologischen Modell der autonomen Regulation basiert.
Jüngste Erkenntnisse stärken die empirische Grundlage der Polyvagal-Theorie. So haben beispielsweise Strain et al. (2024) mittels optogenetischer Aktivierung gezeigt, dass Neuronen im DMNX – einer Schlüsselkomponente des dorsalen Vaguskomplexes – Bradykardie erzeugen und angstähnliches Verhalten modulieren, ohne eine Unterdrückung der Bewegungsfähigkeit auszulösen. Diese Unterscheidung steht im Einklang mit der Differenzierung der Polyvagal-Theorie zwischen dorsal vermittelten Bedrohungsreaktionen und Immobilisierung ohne Angst und räumt mit langjährigen Missverständnissen auf, darunter auch solchen, die in der von Grossman herausgegebenen
Sonderausgabe der Biological Psychology perpetuiert wurden.
Dieser Zusammenhang wirft weitere ethische Bedenken auf. Die von Gastredakteuren betreute Ausgabe enthielt mehrere Kritiken an der Polyvagal-Theorie – etwa von Neuhuber und Berthoud (2022) sowie Doody et al. (2023) –, ohne Gegenargumente oder wissenschaftliche Stellungnahmen anzubieten. Angesichts der bekannten Ablehnung der Polyvagal-Theorie durch Grossman führt das Fehlen gegenteiliger Ansichten zu einer potenziellen redaktionellen Schlagseite. Eine solche Asymmetrie birgt die Gefahr, dass der wissenschaftliche Konsens falsch dargestellt und im akademischen wie auch im öffentlichen Bereich Irrtümer weiterverbreitet werden.
Wie Porges (2025b) feststellt, sollte sich wissenschaftliche Kritik mit Theorien so auseinandersetzen, wie sie verfasst wurden – und nicht mit Karikaturen davon. Ein ethischer wissenschaftlicher Diskurs erfordert Treue gegenüber den Beweisen, Aktualität in der Interpretation und Transparenz in der Methodik. Diese Standards sind nicht nur für eine genaue Bewertung der Polyvagal-Theorie unerlässlich, sondern auch für die Wahrung der Integrität des
wissenschaftlichen Austauschs im umfassenderen Sinne.
Falsche Darstellung im öffentlichen Diskurs
Einige Kritikpunkte, die häufig mit der Polyvagal-Theorie in Verbindung gebracht werden – etwa eine zu starke Vereinfachung oder übermäßige Kommerzialisierung – sind in Wirklichkeit Kritik daran, wie die Theorie von Dritten dargestellt oder angewendet wird, und keine Mängel, die in der Theorie selbst stecken.
- Übermäßige Vereinfachung: Behauptungen, dass die Polyvagal- Theorie die Beziehungen zwischen autonomem Nervensystem und Emotionen über Gebühr vereinfache, beruhen oft auf sekundären Interpretationen, die ihre tieferen neurophysiologischen Dimensionen außer Acht lassen. Konstrukte wie die „Vagusbremse” und „Neurozeption” werden, die durchaus empirisch belegt sind, werden in der öffentlichen Diskussion häufig darauf reduziert, dass es sich um Metaphern handle. In diesen Vereinfachungen spiegelt sich eine mangelnde Auseinandersetzung mit der Primärliteratur; sie sind keine intrinsischen Merkmale der Polyvagal-Theorie.
- Überkommerzialisierung: Es finden sich Adaptionen der Polyvagal-Theorie im Kontext von Coaching, Wellness und Branding, wo es an wissenschaftlicher Stringenz mangeln mag. Zwar lässt dieser Trend berechtigte Bedenken hinsichtlich der translationalen Ethik aufkommen, die Theorie als solche berücksichtigt jedoch ethische Erwägungen durchaus und sollte nicht für Verzerrungen verantwortlich gemacht werden, die auf das Konto externer Akteure gehen.
- Kritik an der anatomischen Unterscheidung zwischen NAmb und DMNX: Neuhuber und Berthoud (2022) argumentieren, dass es für die von der Polyvagal-Theorie postulierte anatomische Trennung keine empirische Grundlage gäbe. Ihre Kritik übersieht jedoch solide Belege für eine Unterscheidung zwischen NAmb und DMNX wie etwa ihre embryonalen Ursprünge, die efferente Myelinisierung und ihre funktionelle Spezifität. Transkriptomische Daten (Coverdell et al., 2019; Jalil et al., 2023) zeigen unverwechselbare molekulare Marker für NAmb-Neuronen, die mit der autonomen Funktion und Myelinisierung im Zusammenhang stehen. Strain et al. (2024) wiesen darüber hinaus nach, dass die selektive optogenetische Aktivierung von DMNX-Neuronen Bradykardie induziert und ein angstähnliches Verhalten moduliert, was die in der Polyvagal-Theorie beschriebene funktionelle und verhaltensbezogene Relevanz des dorsalen Vaguskomplexes bestätigt.
- Phylogenetische Kritik auf der Grundlage nicht säugetierspezifischer Vagusstrukturen: Einige Kritiker berufen sich auf vagale Merkmale bei Lungenfischen oder Reptilien, mit denen sie gegen die Behauptungen der Polyvagal-Theorie zur Evolution zu Felde rücken. Diese Argumente vermischen funktionelle Analogie mit phylogenetischer Homologie. Die Polyvagal-Theorie behauptet nicht, dass die vagale Regulation ausschließlich bei Säugetieren vorkäme, sondern identifiziert den NAmb-vermittelten, myelinisierten Vagus, der in das kraniale soziale Nervensystem integriert ist, als eine Anpassungsleistung der Säugetiere. Die Gleichsetzung der artübergreifenden Vagusfunktion mit diesem spezialisierten System spiegelt ein Missverständnis der evolutionären Spezifität der Polyvagal-Theorie wider.
Es ist wichtig, zwischen der empirischen Grundlage einer Theorie und ihrer öffentlichen Darstellung zu unterscheiden. Der Wikipedia- Eintrag zur Polyvagal-Theorie veranschaulicht diese Vermischung, indem er Missbrauch oder Falschdarstellungen durch Dritte oft so behandelt, als würden sie Mängel in der Theorie selbst offenbaren. Dies stellt einen Kategorienfehler in der wissenschaftlichen Bewertung dar.
Bei genauer Betrachtung zeigen diese Kritikpunkte keine theoretischen Schwächen, sondern Herausforderungen bei der korrekten Verbreitung. Die Wahrung der wissenschaftlichen Integrität der Polyvagal-Theorie erfordert eine präzise Kommunikation und ethisch verantwortliches Handeln von Seiten derjenigen, die sie auf klinischem, edukativem und öffentlichem Gebiet anwenden.
Typologie der Fehler in der Kritik an der Polyvagal-Theorie
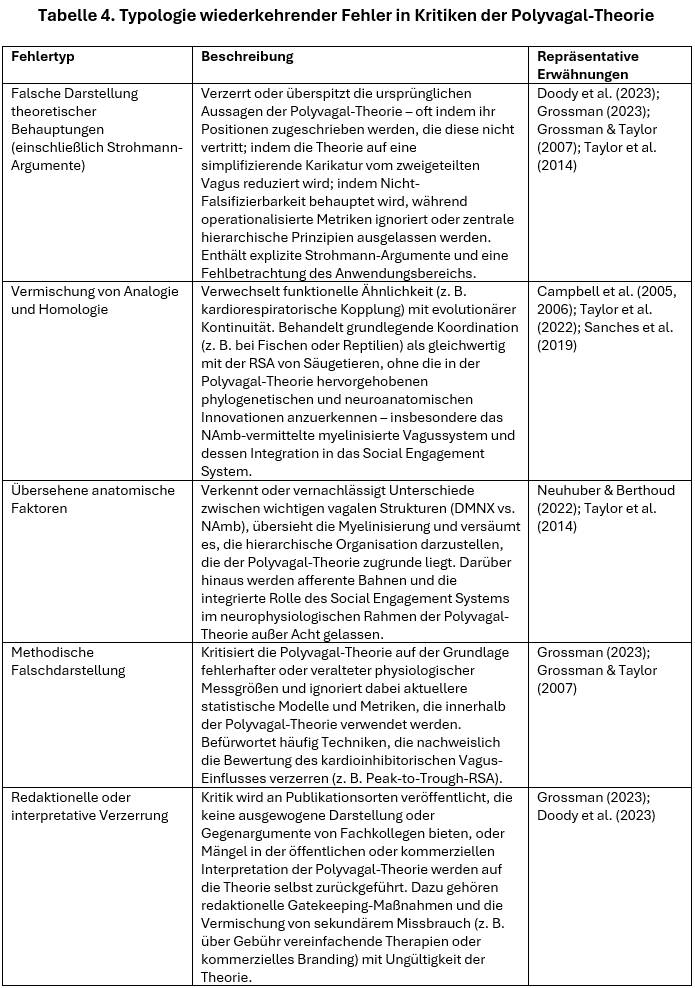
Um die wiederkehrenden Probleme zusammenzufassen, die in der Kritik an der Polyvagal-Theorie identifiziert wurden, präsentiert Tabelle 4 eine Typologie häufiger Fehlerkategorien. Diese reichen von rhetorischen Falschdarstellungen bis hin zu empirischen und methodischen Fehltritten. Jeder Fehlertyp wird definiert, anhand seines Auftretens in veröffentlichten Kritiken veranschaulicht und zu der daraus resultierenden wissenschaftlichen Verzerrung in Verbindung gesetzt.
Diese diagnostische Klassifizierung unterstreicht ein zentrales Anliegen des vorliegenden Dokuments: Die hartnäckigsten und sichtbarsten Kritikpunkte an der Polyvagal-Theorie setzen sich oft nicht mit den empirischen und konzeptionellen Aspekten der Theorie auseinander. Anstatt fundierte wissenschaftliche Einwände vorzubringen, spiegeln diese Kritiken häufig Missverständnisse imó Hinblick auf die Evolutionsbiologie, Neuroanatomie und methodischen Validität wider – oder entstehen aus redaktionellen und translationalen Dynamiken, die jenseits der Theorie selbst angesiedelt sind.
Fazit
Eine umfassende Auswertung der veröffentlichten Kritik an der Polyvagal-Theorie zeigt eine wiederkehrende Fehlerstruktur: Die einflussreichsten Einwände – auf anatomischem, physiologischem, evolutionärem und konzeptionellen Gebiet – lassen sich immer wieder auf die wissenschaftlichen Beiträge und den redaktionellen Einfluss von Paul Grossman zurückführen. Ob direkt durch von ihm verfasste Artikel, durch Zusammenarbeit mit E. W. Taylor oder die redaktionelle Kuratierung der Sonderausgabe von Biological Psychology aus dem Jahr 2023 – Grossman hat eine zentrale Rolle bei der Gestaltung des öffentlichen und akademischen Diskurses um die Polyvagal-Theorie gespielt.
Diese Konzentration von Dissens, insbesondere wenn sie durch Publikationsorte verstärkt wird, die keine Gegendarstellung zulassen, wirft berechtigte Bedenken in Sachen wissenschaftliche Objektivität und Vortäuschung von Konsens auf. Wissenschaftliche
Kritik muss auf einer zutreffenden Darstellung der Grundaussagen einer Theorie beruhen, nicht auf einer Karikatur hiervon oder auf rhetorischen Ablenkungsmanövern. Wie dieses Manuskript gezeigt hat, stellen viele Kritiken der Polyvagal-Theorie deren Kernkonzepte falsch dar, oft mittels einer kleinen Anzahl von kategorischen Fehlern, die sich in verschiedenen Publikationen und Plattformen wiederholen.
Eine solche Wiederholung sollte nicht mit Konvergenz oder Konsens verwechselt werden. Vielmehr unterstreicht sie die Notwendigkeit einer größeren epistemischen Verantwortlichkeit sowohl im wissenschaftlichen als auch im öffentlichen Bereich. Leser und Gutachter werden dringend gebeten, direkt die Primärliteratur heranzuziehen, in der die empirischen Grundlagen und die konzeptionelle Architektur der Polyvagal-Theorie am deutlichsten dargelegt sind.
Hier finden Sie die aktuellsten wissenschaftlichen Artikel von Dr. Stephen Porges:
Februar 2026: When a Critique Becomes Untenable: A Scholarly Response to Grossman et al.’s Evaluation of Polyvagal Theory
September 2025: Polyvagal theory: a journey from physiological observation to neural innervation and clinical insight
Sie möchten mehr Informationen zum Vagusnerv?
Quellennachweise
Byrne, E. A., & Porges, S. W. (1993). Data-dependent peak–valley estimation of respiratory sinus arrhythmia: A methodological evaluation. Psychophysiology, 30(6), 672–678. https://doi.org/10.1111/j.1469-8986.1993.tb02061.x
Campbell, H. A., Taylor, E. W., & Egginton, S. (2005). Does respiratory sinus arrhythmia occur in fishes? Biology Letters, 1(4), 484–487. https://doi.org/10.1098/rsbl.2005.0365
Anmerkung: Vermischung von Analogie und Homologie – Behandelt die kardiorespiratorische Kopplung bei Fischen als homolog zur RSA bei Säugetieren, verwechselt dabei funktionelle Ähnlichkeit mit phylogenetischer Kontinuität und übersieht die NAmb-basierte, myelinisierte Spezialisierung bei Säugetieren, die für die Polyvagal-Theorie von zentraler Bedeutung ist.
Campbell, H. A., Leite, C. A. C., Wang, T., et al. (2006). Evidence for a respiratory component… in the rattlesnake. Journal of Experimental Biology, 209(14), 2628–2636. https://doi.org/10.1242/jeb.02278
Anmerkung: Vermischung von Analogie und Homologie – Verwendet reptilische RSA-ähnliche Kopplung, um eine evolutionäre Äquivalenz mit der RSA von Säugetieren zu implizieren, und umgeht dabei die Betonung der NAmb- Innovationen und SES-Integration bei Säugetieren im Rahmen der Polyvagal-Theorie.
Coverdell, T., Ivison, R., Tao, J. und Campbell, J. (2019). 582-P: Disambiguating the nucleus ambiguus with single-cell transcriptomics. Diabetes 68 (Suppl. 1):582. https://doi.org/10.2337/db19-582-P
Dale, L. P., Kolacz, J., Mazmanyan, J., Leon, K. G., Johonnot, K., Bossemeyer Biernacki, N., & Porges, S. W. (2022). Childhood maltreatment influences autonomic regulation and mental health in college students. Frontiers in Psychiatry, 13, 841749. https://doi.org/10.3389/fpsyt.2022.841749
Doody, J. S., Burghardt, G. M., & Dinets, V. (2023). The evolution of sociality and the polyvagal theory. Biological Psychology, 180, Article 108569. https://doi.org/10.1016/j.biopsycho.2023.108569
Anmerkung: Falsche Darstellung theoretischer Behauptungen (einschließlich Strohmann-Argumente) – Die Polyvagal- Theorie wird so umgedeutet, als behaupte sie, Reptilien seien „unsozial”, und das Argument der Theorie, dass Säugetiere über eine spezifische autonome Innovation verfügen, wird als Leugnung der Sozialität von Reptilien behandelt, wobei Verhaltensanalogien mit dem neuroanatomischen Betrachtungsrahmen der Polyvagal-Theorie verwechselt werden.
Grossman, P. (2023). Fundamental challenges and likely refutations of the five basic premises of the polyvagal theory. Biological Psychology, 180, 108589. https://doi.org/10.1016/j.biopsycho.2023.108589
Hinweis: Falsche Darstellung theoretischer Behauptungen (Strohmann-Argument) – Der Polyvagal-Theorie werden Behauptungen zugeschrieben, die sie nicht aufstellt (z. B. die ausschließliche Gültigkeit der RSA für Säugetiere). Falsche Darstellung der anatomischen Spezifität und physiologischen Nuancen der Theorie. Nichtberücksichtigung der klar unterscheidbaren kardioinhibitorischen myelinisierten Nervenbahnen des Nucleus ambiguus (NAmb) und ihrer Integration in die kranialen motorischen Systeme, die das Social Engagement System bilden, sowie der koordinierenden Rolle des gemeinsamen kardiopulmonalen Oszillators bei der autonomen Regulation von Säugetieren.
Grossman, P., & Taylor, E. W. (2007). Toward understanding respiratory sinus arrhythmia: Relations to cardiac vagal tone, evolution and biobehavioral functions. Biological Psychology, 74(2), 263–285. https://doi.org/10.1016/j.biopsycho.2005.11.014
Anmerkung: Falsche Darstellung von Behauptungen der Theorie (einschließlich Strohmann-Argumente) – Baut einen Strohmann im Hinblick auf die evolutions- und vagusbezogenen Aussagen der Polyvagal-Theorie auf, zeigt Anleihen bei Konstrukten der Polyvagal-Theorie ohne korrekte Nennung des Urhebers und hebt gestützt auf ein nichtiges RSA-Framing die Unterscheidung zwischen NAmb und DMNX auf.
Jalil, M., Coverdell, T. C., Gutierrez, V. A., Crook, M. E., Shi, J., Stornetta, D. S., Schwalbe, D. C., Abbott, S. B. G., & Campbell, J. N. (2023). Molecular disambiguation of heart rate control by the nucleus ambiguus [Vorabdruck]. bioRxiv. https://doi.org/10.1101/2023.12.16.571991
Kolacz, J., Roath, O. K., Lewis, G. F., Karrento, K., et al. (2025). Cardiac vagal efficiency is enhanced by percutaneous auricular neurostimulation in adolescents with nausea: Moderation by antidepressant drug exposure. Neurogastroenterology & Motility, 37(5), e15007. https://doi.org/10.1111/nmo.15007
Lewis, G. F., Furman, S. A., McCool, M. F., & Porges, S. W. (2012). Statistical strategies to quantify respiratory sinus arrhythmia: A comparative evaluation. Biological Psychology, 89(2), 349–364. https://doi.org/10.1016/j.biopsycho.2011.11.009
Monteiro, D. A., Taylor, E. W., Sartori, M. R., Cruz, A. L., Rantin, F. T., & Leite, C. A. (2018). Cardiorespiratory interactions previously identified as mammalian are present in the primitive lungfish. Science Advances, 4(2), eaaq0800. https://doi.org/10.1126/sciadv.aaq0800
Anmerkung: Vermischung von Analogie und Homologie – Leitet aus funktionellen Ähnlichkeiten eine evolutionäre Homologie bei den kardiorespiratorischen Interaktionen von Lungenfischen ab und übersieht dabei den Fokus der Polyvagal-Theorie auf die säugetierspezifische’ neuroanatomische Reorganisation, insbesondere die mit dem NAmb zusammenhängende Myelinisierung und die Integration mit dem Social Engagement System als evolutionäre Innovationen.
Neuhuber, W. L., & Berthoud, H.-R. (2022). Functional anatomy of the vagus system: How does the polyvagal theory comply? Biological Psychology, 174, 108425. https://doi.org/10.1016/j.biopsycho.2022.108425
Anmerkung: Anatomische Unachtsamkeit – Reduziert die Polyvagal-Theorie auf ein unzulässig vereinfachendes Narrativ vom „Doppel-Vagus”, während sie die Unterschiede in der Entwicklung, Myelinisierung und Hierarchie zwischen NAmb und DMNX umgeht. Ignoriert afferente Bahnen und lässt das Social Engagement System außer Acht, wodurch die schwerpunktmäßige Beschäftigung mit integrierten funktionellen Schaltkreisen und evolutionärer Organisation in der Polyvagal-Theorie übersehen wird.
Porges, S. W. (1995). Orienting in a defensive world: Mammalian modifications of our evolutionary heritage: A Polyvagal Theory. Psychophysiology, 32(4), 301–318. https://doi.org/10.1111/j.1469- 8986.1995.tb01213.x
Porges, S. W. (1998). Love: An emergent property of the mammalian autonomic nervous system. Psychoneuroendocrinology, 23(8), 837–861. https://doi.org/10.1016/S0306-4530(98)00057-2
Porges, S. W. (2003). Social engagement and attachment: A phylogenetic perspective. Annals of the New York Academy of Sciences, 1008(1), 31–47. https://doi.org/10.1196/annals.1301.004
Porges, S. W. (2004). Neuroception: A subconscious system for detecting threats and safety. Zero to Three, 24(5), 19–24.
Porges, S. W. (2007a). The polyvagal perspective. Biological Psychology, 74(2), 116–143. https://doi.org/10.1016/j.biopsycho.2006.06.009
Porges, S. W. (2007b). A phylogenetic journey through the vague and ambiguous Xth cranial nerve: A commentary on contemporary heart rate variability research. Biological Psychology, 74(2), 301- 307. https://doi.org/10.1016/j.biopsycho.2006.08.007
Porges, S. W. (2022). Polyvagal theory: A science of safety. Frontiers in Integrative Neuroscience, 16, 871227. https://doi.org/10.3389/fnint.2022.871227
Porges, S. W. (2023). The vagal paradox: A polyvagal solution. Comprehensive Psychoneuroendocrinology, 16, 100200. https://doi.org/10.1016/j.cpnec.2023.100200
Porges, S. W. (2025a). Polyvagal Theory: Current status, clinical applications, and future directions. Clinical Neuropsychiatry, 22(1), 7–26. https://doi.org/10.36131/cnfioritieditore20250301
Porges, S. W. (2025b). Polyvagal theory: a journey from physiological observation to neural innervation and clinical insight. Frontiers in Behavioral Neuroscience, 19, 1659083. https://doi.org/10.3389/fnbeh.2025.1659083
Porges, S. W., & Furman, S. A. (2011). The early development of the autonomic nervous system provides a neural platform for social behavior: A Polyvagal Perspective. Infant and Child Development, 20(1), 106–118. https://doi.org/10.1002/icd.688
Reed, S. F., Ohel, G., David, R., & Porges, S. W. (1999). A neural explanation of fetal heart rate patterns: A test of the Polyvagal Theory. Developmental Psychobiology, 35(2), 108–118. https://doi.org/10.1002/(SICI)1098- 2302(199909)35:2%3C108::AID-DEV4%3E3.0.CO;2-N
Richter, D. W., & Spyer, K. M. (1990). Cardiorespiratory control. In A. D. Loewy & K. M. Spyer (Eds.), Central regulation of autonomic functions (pp. 189–207). Oxford University Press. https://doi.org/10.1093/oso/9780195051063.003.0011
Sanches, P. V., Taylor, E. W., Durán, L. M., Cruz, A. L., Dias, D. P., & Leite, C. A. C. (2019). Respiratory sinus arrhythmia is a major component of heart rate variability in undisturbed, remotely monitored rattlesnakes (Crotalus durissus). Journal of Experimental Biology, 222, jeb197954. https://doi.org/10.1242/jeb.197954
Anmerkung: Vermischung von Analogie und Homologie – Bezeichnet die reptilische Kopplung als RSA, die der NAmb- vermittelten RSA von Säugetieren entspräche, und ignoriert dabei die säugetierspezifische neuroanatomische und verhaltensbezogene Integration in der PVT.
Strain, M. M., Conley, N. J., Kauffman, L. S., Espinoza, L., Fedorchak, S., Castro Martinez, P., … Boychuk, C. R. (2024). Dorsal motor vagal neurons can elicit bradycardia and reduce anxiety-like behavior. iScience, 27(3), 109137. https://doi.org/10.1016/j.isci.2024.109137
Taylor, E. W., Campbell, H. A., Levings, J. J., Young, M. J., Butler, P. J., & Egginton, S. (2006). Coupling of the respiratory rhythm in fish with activity in hypobranchial nerves and with heartbeat. Physiological and Biochemical Zoology, 79(6), 1000–1009. https://doi.org/10.1086/507663
Anmerkung: Vermischung von Analogie und Homologie – Interpretiert die Kopplung von Atmung und Herzfrequenz bei Fischen als Beweis für die evolutionäre Kontinuität bezogen auf die respiratorische Sinusarrhythmie bei Säugetieren, ohne auf die säugetierspezifischen mit dem NAmb verknüpften, myelinisierten Vagusbahnen und ihre Integration in das Social Engagement System einzugehen, die im Mittelpunkt der Polyvagal-Theorie stehen.
Taylor, E. W., Campbell, H. A., Levings, J. J., Young, M. J., Butler,. J., & Egginton, S. (2006). Coupling of the respiratory rhythm in fish with activity in hypobranchial nerves and with heartbeat. Physiological and Biochemical Zoology, 79(6), 1000–1009. https://doi.org/10.1086/507663
Anmerkung: Falsche Darstellung theoretischer Behauptungen (einschließlich Strohmann-Argumente) – Deutet die Polyvagal-Theorie dahingehend um, dass diese die Exklusivität der kardiorespiratorischen Kopplung bei Säugetieren postuliere und kritisiert dann diese umformulierte Behauptung, anstatt sich mit der in der Theorie betonten neuroanatomischen Spezialisierung bei Säugetieren auseinanderzusetzen, insbesondere der Integration des ventrovagalen Komplexes, der soziale Interaktionen unterstützt.
Copyright 2026, Stephen W. Porges